|
RESONANCE & COHERENCE
From harmony,
from heavenly harmony,
This universal frame began:
From harmony
to harmony
Through all the compass of the notes it ran, The diapason closing full on man.
Dryden in Trusted (1991)
|
|
| The chromosome is strung across the nucleus from the nucleolus to the nuclear membrane |
Having now established the means
by which material and energetic standing wave phenomena inform the genome, there is some substantiation to this modelling
to be found by furthering our reconstruction of the half arm set we have taken as fundamental. As was earlier explained,
the modelling has been based on the Alberts et al., graphic adapted from the data of Ute Franke et al., and we can return
to this to try and find some means of illustrating the harmonic resonances that underly the informational principles outlined.
Perhaps first is required a brief explanation of how the modelling is done, childishly simple though it is.
The set graphic is photocopied onto transparency, the chromosomes cut out, cut across the waist of the chromosome and, hey
presto, we play! Dividing down the half arm set in similar fashion to that previously used in the double structure set
to illustrate the self-assorting of the five driver arms, we can then take the 8 or 9 resulting half arms that constitute
each sub-set and overlay them, using the characteristic banding pattern of the giemsa staining as an informational indicator.
It is rather binary, the black and white of the stained array and we will later address the issue of what the staining actually
tells us, but for now it is sufficient to accept that it does work as a yes/no, on/off etc., type of pattern information,
essentially indicating a two state system distributed through the half chromosome set members of the genome.
When
each of the harmonic sub-sets is overlaid - and, if you do actually do this you cannot see what is happening until your run
the constructed sets through a photocopier again - we can first define what we might expect to see. Given that the seb-set
members are here proposed as forming a closed set within themselves, we might expect that in overlay the staining would give
rise to an aggregate object that is completely committed to one state and, obviously this would result in a totally black
or white object. Of course by this method we cannot generate a totally white sub-set object and must work with the dark
giema stain as indicator. When we do construct the sets in overlay we find that in fact the staining is not complete
and a clearly defined 'resonance window' appears within each of the five sub-sets.
|
|
| Each half arm sub-set produces a resonance window |
As will be seen in the CODA sub-page to
this the staining in fact can be taken as a general indicator of the dominant base sequence of the mapped chromosome arm.
Indeed, the Alberts et al., legend itself tells us that the giemsa stain preferentially binds A-T rich base sequences and
the non-stained regions we can therefore take as predominantly C-G rich. This is indicator of what in CODA become defined
as the 'evolved' amino acids and these are then to be understood as 'evolutionary' windows. The A-T
rich regions, then, we take as indicator of basal metabolism on which any evolutionary experiment and development must necessarily
depend.
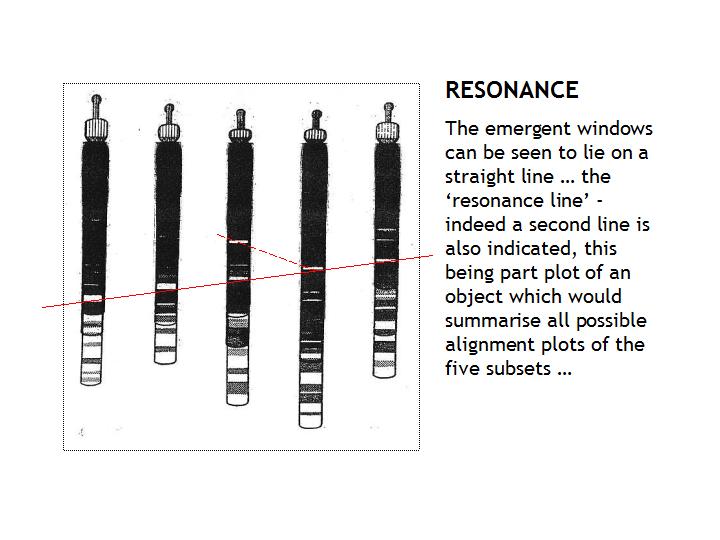
Whilst each window within the sub-sets indicates the resonance between the set members, when
the five sets are laid out and ordered by resonance logic we see, as illustrated below, that the windows can be set out to
form a resonance line across the five sub-sets and this we can fairly deduce is indicative of the whole set achieving a resonance
of resonances - in other words, whole set coherent entrainment.
The fact that we can do this at all, get any pattern
whatsoever, despite three levels of error, tells us something reasonable and robust must be buried in all this or it is an
erroneous construction only realised by extraordinary fortuosity. I of course prefer the former interpretation!
|
|
| A resonance line links the five 'evolutionary windows' |
In the above illustration
the five parallel processing sub-sets of the genome are laid out in an order whereby the 'resonance' windows are
formed into a straight line, the 'resonance of resonances' which we take to indicate coherence within the set.
As can be seen also, a second and only partial line is indicated, its continuance being buried within other sub groups of
each of the sub-sets.
This single deconstruction has all been made by reference to the chromosome 'waist'
and we can go on to predict then that, given the robustness that must exist for this pattern to emerge at
all, and the system robustness that we know in truth underlies it, a similar resonance line will be generated when we change
our modelling to take, for example, the centromere edge or the telomere as our fixed reference point. Indeed, taking
any isographic point as reference across the sets, we must suspect that on each occasion a resonance line and partial second
line will always emerge.
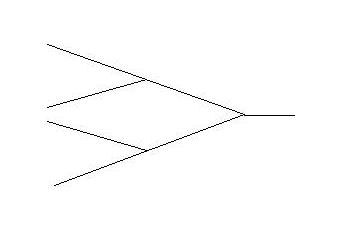
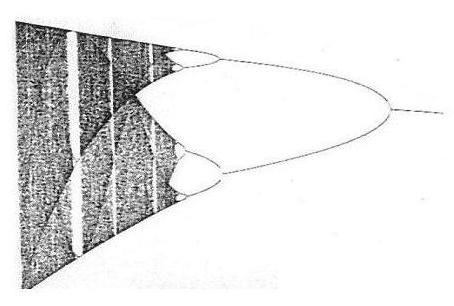
When in abstract we generate a summary object of all these arrangements we will
of course end up with the now familiar and iconographic bifurcation diagram as indicator of the true dynamic object we are
heading for which is, of course, the Feigenbaum.
We are then by this construction invited
to consider what it means that man's genome can indeed be understood as a quantum system and at the
foundation of which we find clear signature of chaos? What then might be said of the profundity of the mathematic
and, yet, are we really surprised to find chaos as the fundamental embedding of man's own genome? Do we not,
in truth, somehow already know, by virtue of the mathematics of fractal geometry and chaos theory that we are doing today
and our ever-increasing understanding of complex systems, that man must indeed stand - quite literally - and have the ground
of his own being as some sort of answer to the chaos that is the quantum realm?
These are today, at one and the same time, both
practical and philosophical questions, such is the powerful scope of the new mathematical schema that is both metric and conceptual
framework.
In terms of the practical reality - that the telomere effectively constitutes a magnetic ion trap qubit
- there is initially great discomfort (for some) in the suggestion, yet on reflection one has to accept that this mainly
arises by the constraint of imagination that has formerly limited our thinking on practical computing to silicon and
'machines'.
A rapid review of scientific interests, however, quickly illustrates that our imagination
needs update when we find that at the forefront of 'unconventional computing' are researches around computing
with liquids and with light (see SPUC - Science and Philosophy of Unconventional Computing website) and, then too,
at the forefront of quantum life science exist researches to do with biosystems' energetic dependence on light-based or
photonic relay in the form of 'biophotons' (see the work of Fritz Albert Popp, IIB website).
Similarly,
as a necessary strategic move in hope of evading Moore's computational limit, the computing industry has
itself in review and is today moving away from silicon as the material substrate of computing and exploring the computational
possibilities and capabilities of the various forms of carbon - buckyballs, buckytubes, graphene - as the new substrate and
'machinery' of computation.
The co-mapping of these interest sets and characterisation of their
focus as 'soft systems' at least offers the means of our beginning upon realising the power and extent of
quantum self-organisation and our being and consciousness as product.
Science rightly progresses by holding
all current scientific 'truths' as provisional, history of course having taught that what today is perceived to stand
as 'truth' is tomorrow felled by discovery. Thus it is that the foundations of quantum theory
are still held as incomplete and, so too then, the full elaboration of what quantum theory might offer is necessarily very
incomplete. We do not yet know the limits or bounds of quantum systems in either the most simple of circumstances or,
as we find here in living systems, the most highly elaborated and coherently self-organised of systems built around exploiting
the range of energies released by natural - physical - process and the extension of this range by self recreation (internal
generation) of far-from-equilibrium conditions, e.g., the output product of 'sloppy' protein working
is crystalline DNA; the variety of the essential amino acids is replicated in the essential qualities of the 'evolved'
amino acids, whilst the varietal range is then also extended by other of the 'evolved' amino acids, themselves then
the exemplar acidic amino acids.
The possibility that has been illustrated
here - that the g-quadruplex sequence that dominates the telomere might have functional identity as a qubit of the magnetic
ion trap (the ponderomotive, I like to think) variety - comes as a shocking reminder of the power of nature, that quantum
computing should be literally realised in the emergence of organic nature - living systems - and that man, late product of
such, should so lately realise it. Whilst man the scientist has invented various vast, elaborate and very expensive
means to explore our understanding of the atom, living systems, as vast quantum connected atomic systems, must necessarily
already know more than we can imagine about the nature of atoms and the nature of quanta. Perhaps also this is indication
that our discoveries and inventions all have ontological priority in the cell; our creativity is organically informed?
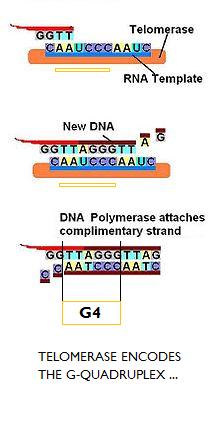
That the g-quadruplex is the primary
qubit is a proposal incidentally evidenced by the fact that the telomerase moiety, building the telomeres, carries
an RNA template whose sequence, as any number of graphics pulled up on the web illustrate, is that of the g-quadruplex,
but which then in the chromosomal DNA is formed in two parts.
As can be readily appreciated, in light
of the possibility that has been raised by our gedanken experiment, quite a number of areas of our current thinking
are thrown into different relief and out of this a 'new' pattern emerges as underpinning the conceptual
development of future science.
A whole new science is opening up ...
|