|
THE PHYSICAL CELL:
Having speculated as to the quantum arrangements that
might exist in the cellular membrane systems and skeletal elements, we now turn to the contents of the cell lumen. The
above is intended to illustrate the nature of the energetic forces acting to ordinate the cell as an environment and within
which the genomic and protein process behaviours play.
Primary in our consideration is the membrane
potential and the field across the cell membrane which is 'of the order of 107 V/m. This field is strong enough
to align all the macromolecules within a biological membrane relative to this field direction, thereby increasing their mutual
interactions and their non-linear dielectric responses to external electromagnetic fields' (Smith, 1989). This field
strength, also then, will have concomitant effect for the energetics of membrane bound proteins which are, effectively, dielectric
dipoles, and, given their functional role as signal transducers and amplifiers within and across the membrane, must also then
inform the internal state of the cell. Thus, given the bounded state of the cell, this would result in a phenomenal
concentration of power and considering what happens to bulk water under high voltage (picture below) we have to suspect that
for cell contained water the state of high potential, tension-driven, self-organisation will also be extraordinarily high.
Indeed, Dubrov (1978) quotes Trincher as stating that 'water inside the cell is in a state of maximum order - in a state
attainable in non-living systems only at absolute zero.' (from Smith, 1989).
Within
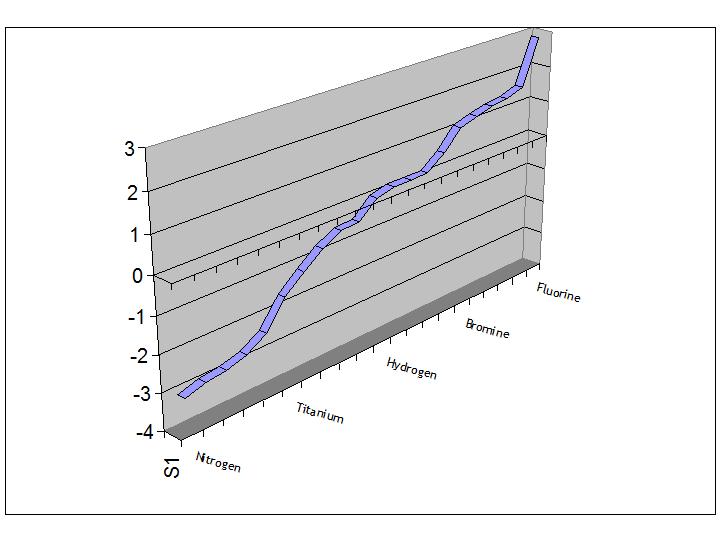
this highly ordered array we have to wonder at the nature of the fundamental electrolyte in terms of living systems.
When we inspect the array of electrode potentials of the atoms used in living systems, for instance, we notice that whereas
general theory might allow that the biochemistry of life might be based around a beginning in the natural saline of the sea and
so might have continued in the quantum scale set by sodium and chorine, living systems seem to have extended the quantum
environment capabilities to work with the more powerful cations of calcium and, ultimately, potassium, whilst
the anionic power ranges beyond chlorine to its quantum cousin fluorine.
|
| The electrolyte as a reaction space - plot of atomic electrode potentials |
From here we move to consideration of the main cellular components in terms of energetic
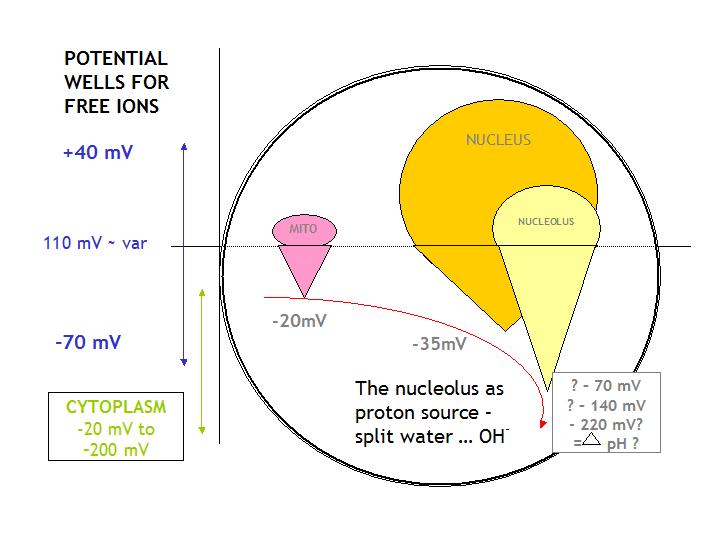
processing, the mitochondria and the nucleus. Taking the cytosol as pH 7 Ling (1984) found potential changes of ~-20mV
for the mitochondria and ~-35mV for the nucleus, making a stepwise progression of potential wells and from which we are invited
to extrapolate the nucleolar pH/potential. We would expect this to present a much stronger potential well than the nucleus,
being itself the caustic focus emergent within the potential ground of the nucleus.
There is a weird
hypersystems anomaly that crops up here. Defininitions of life often talk about respiration, either organismal scale
or molecular, but here in the cell we know by the very nature of the end product of the process that free oxygen is not at
all times welcome when DNA is the system in focus. That is why the end product is deoxyribose and why the cell symbiont
that thrived and evolved into the mitochondrion did so by dint of having discovered a guaranteed oxygen rich environment,
mopping up all the free oxygen which might otherwise corrupt the DNA coding.
Free oxygen in fact acts by causing
the switch of base pairs from GC to AT at the next copying. If this mechanism was found to be operating for AT
to GC transition also, there would be a good model to be constructed of an effectively respirational ratchet system shifting
the code base along one step every copying, 'breath' or cell division. Certainly this offers a Turing
model of sorts. Is there perhaps a reverse mechanism or 'expiration' then that switches all the code
back we might ask?
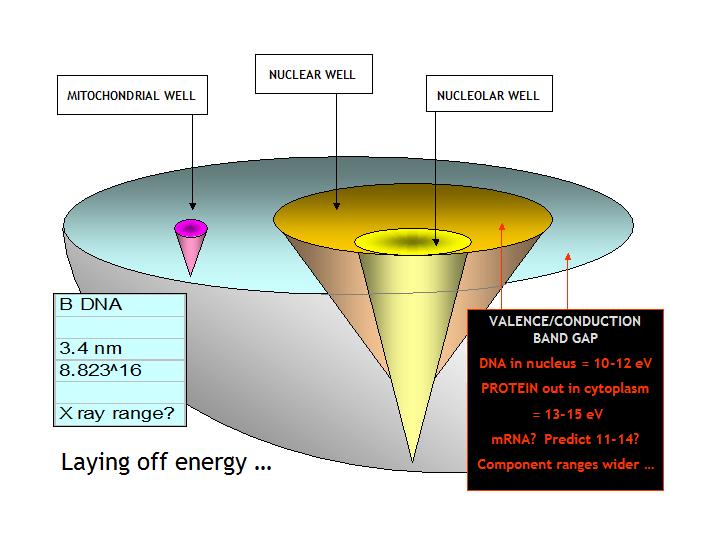
The above illustration invites address of the issue of the
cellular environment in terms of some of the main biological players: the mitochondrial sub-population, nuclear or genome
dynamics and the processing of nuclear output in terms of both the DNA being transcribed within the nucleus and the driven
dispersion of the derived mRNAs - the 'transcriptome' - across the cell interior. It is noticeable that
the concentric arrangement of the array in terms of band gap or activation energies (from Bakhshi, 1994) shows the nuclear
DNA to have an extraordinarily narrow bandgap or low activation energy, whilst the driven population of cytoplasmic mRNA transcripts
and protein translates, respectively, require ever-greater energies. This suggests the nucleus as an energetic
source and the nucleo/cytoplasm as a graduated sink.
In this we also have simple illustration of the transitions
effected by living systems and how this intertwines with some physical understanding of entropy. Given the above data
it has to be noted that the range of activation energies of the individual amino acids is far greater than that of their aggregate,
the protein. Degrees of freedom are lost for the individual in the creation of each and every bond or aggregate, but
in building such aggregates, however, there is the further opportunity for the system to open up new component behaviours
or gain a new type of order as the constituent components in the aggregate are taken beyond their 'comfortable' operational
range. Anabolic processes will reduce entropy and componental degrees of freedom, but by their aggregation of charge
or mass then also acquire properties and freedoms beyond those of any of the individual components.
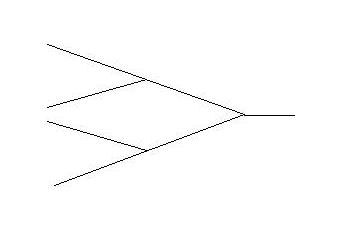
This can be
very simply illustrated by a 'skeletal' Feigenbaum, the iconographic signifier of chaos, which here might be taken
at the 'unique' or 'output' end as the protein and at the 'input' or 'varietal' end as
the amino acid array.
 The preceding illustration of the cell as environment
also seeks to point up the winding frequency of DNA. In B form DNA, regarded as the 'working' pitch of the helix,
i.e., when in transcription to mRNA, the average distance between base pair planes is 0.34nm which, given that the B form
pitch has ten base pairs to the turn and therefore gives a fundamental wavelength of 3.4nm, places the winding of DNA in the
range of X-ray frequencies.
Beyond this there are two further thoughts to bear in mind:
(i) X-rays are capable of producing photon emission, this being the photoelectric effect that Einstein explored
(ii) This is because electron frequencies in the atom are in X-ray range and thus the incidence of a
resonant frequency X-ray photon can excite an electron briefly to a higher orbital and a photon then be relayed to escape
as the electron falls back to the atomic ground state. The frequency of the relay photon must be of longer wavelength
than the incident X-ray photon, energy having been lost to the atomic system in accommodating the photon reception, process
and onward flight.
That the cell is an essentially quantum
system can perhaps more readily be understood by a simple thought experiment based on a sort of reverse engineering whereby,
for example, biological methods use gel electrophoresis as a means of ordinating the object arrays of their interest in 1
or 2 dimensions. Bioelectromagnetics would propose that this too is the mechanism by which the system itself naturally
works - indeed, can only work - and long ago established the currents that flow within the tissues of living
systems. The cell naturally operates as a three dimensional electrophoretic system ordinating naturally polar dielectric
proteins and ions.
This fact in itself can assist in unravelling evolutionary stories in so far as instances of
misplacement within an expected electrophoretic distribution indicates that some contingent history has united elements that
would otherwise have their own different dynamic histories.
From
this general outline then the character of the cellular environment is proposed as that of a cyclically acting, self-electrophoresing
system whose feedback dynamics are naturally focussed at the nucleus. This is on the grounds that although gel electrophoresis
is sufficient for the biologist to achieve control and spatio-temporal order in the sub-cellular array of their interest,
no model has to date been developed taking this as a means of ordination for the cell itself.
> THE PHYSICAL CELL: FLOW
|