|
THE PHYSICAL CELL:

|
| The dynamics of the cell cycle |
|
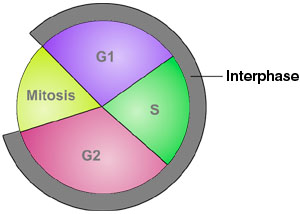
| The cell at 1 o'clock (left) is in interphase state, other figures mitotic |
|
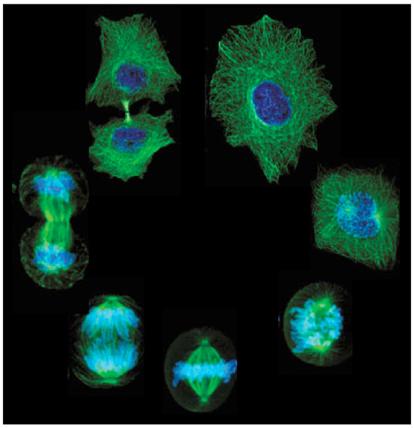
| The array of states in DNA and cell duplication and division |
The above animation of the cell cycle would usually be illustrated by some such as the cell states array (above, middle)
or the phase diagram (above, right), each in their way marking the general sub-periods that have been defined for the
normal process of cellular working and duplication. This can also be represented by a loop within a loop whereby
the interphase state includes the synthetic phase of DNA duplication.
Cellular self-replication,
which in essence relies on the population dynamics of mitochondria (whose reproduction runs at something like 10x the
cellular rate) as these also have function as oxygen attractors, or sinks, extends then from 'birth' of the independent
cell marking the end of the division process (mitosis), through re-establishment of nuclear working (G1) to enable DNA
synthesis (S), until the onset of the next division process (at the end of G2). The cytoplasmic 'duplication'
that is required to produce two cells of similar state to the progenitor thus introduces a third loop of coupled systems.
It is just such threefold systems that can produce chaotic dynamics.
|
|
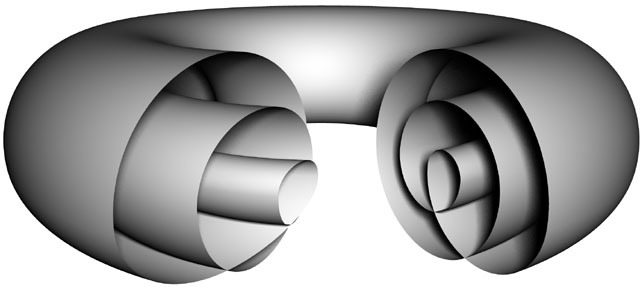
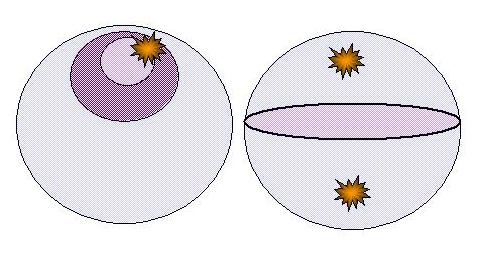
| The triple toroid embedding that can model the cell |
|
|
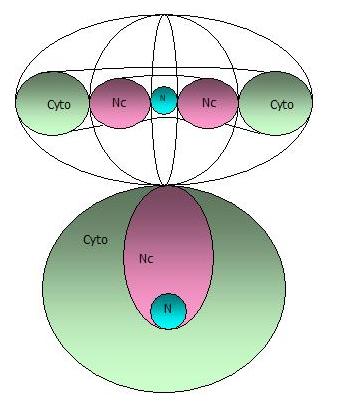
| Diagrammatic representation of the cell as a set of three toroids |
The
toroidal model is chosen because of the vesicular traffic between the exterior of the cell, i.e., the cell environment, and
the input and output flows to the cytoplasm. Modelling likewise for the nucleus and nucleolus respectively embeds each
within the other, but each then also has direct connection to their external environment. Each also models as a caustic
boundary because of that input mode.
The relative timing of events here is very variable between
cell types, if not signature for them, and the duration of each of the cell phases is also highly variable. Whilst this
tells us much about the extraordinary tolerance or robustness of the system, it does not tell us about the real dynamics for
the working cell, i.e., in situ. Of course many of the cells of the body do not replicate at all, i.e., do not cycle,
but are thrown off by their mother cells to a singular fate, e.g., epidermal cells or erythrocytes. It is as yet
unrecognised, but here felt to be significant, that the stem cell populations that throw off these daughter cells are
in most part attached to the basement membrane, which is, as an amplification of the cell membrane model, the Faraday
cage of the multicellular organism. Other stem cell populations within the organism are themselves contained within
further membrane boundaries, e.g., testes, ovaries, spleen.
As elsewhere
on this site, simple illustration is felt to be more readily informative for the non-specialist, so illustration
is given below to the two cellular states which, in the model to be developed here, are taken as the extreme
points of the cycle in terms of cellular geometry and dynamics, these being the 'working' and 'metaphase'
states of the cell.
|
|
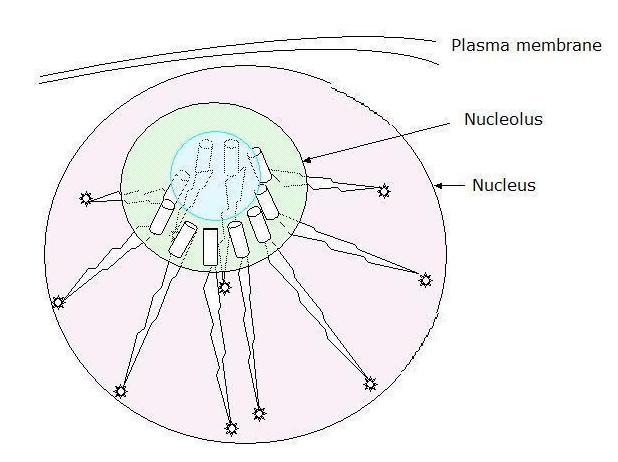
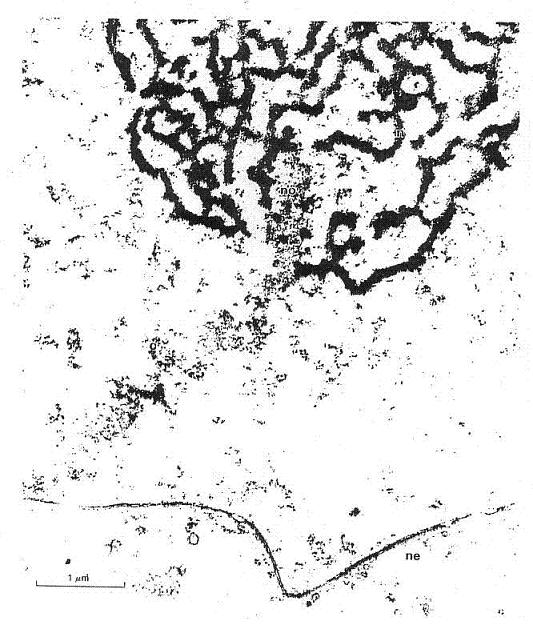
| Diagram of arrangements - co-aggregation of the five (x2) rRNA encoding arms in the nucleolus |
Working
state
The above is intended as diagrammatic illustration of the working state which is here presented
so as to make clear the functional relations of five 'half' chromosomes that are highly condensed, highly iterated
DNA sequences coding for rRNAs carried on chromosomes 13, 14, 15, 21 and 22. We will be addressing their functionality
more closely in the next section.
When not coding for rRNA, i.e., involved in the process of transcription, these
structural arrays can then also be seen as presenting a quasi-periodic crystalline array which, taken in section, could also
act as a photonic - or indeed ionic - crystal.
|
|
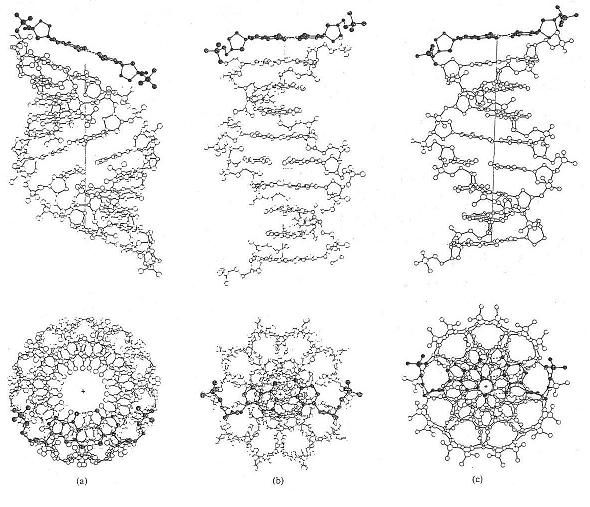
| Core symmetries depend on type/pitch of DNA - A, B and C above, l to r |
Most noticeable is that the form proposed
to be the active DNA state, i.e., the B DNA form, is that point in the pitch twisting between states when the axis of the
DNA core is at 90o to the plane of the base pair bonds across the helix. Given that the bases are resonant
structures, i.e., cyclic, and have strong pi-orbitals, it has to be suspected that there is some possible function
here for the plane of the base-pairs to act as an electromagnetic mirror or 'state splitter'.
It is notable also that in the working
state the general disposition of the nucleolus generating arms is to be strung across from the nucelolar core to the periphery
of the nucleus itself and the picture given below, albeit artifact or otherwise, neatly illustrates the sort of thinking that
the physicists - such as Frohlich and Popp - have in mind when addressing issues around the propagation of waves, 'solitons'
or 'polarons' along the length of the DNA. Here we seem to have captured just such a wave in process. This general
arrangement indicates that we need to look at standing wave phenomena given that these particular half chromosomes or chromosome
arms extend between the geometrically invariant arrangements that give rise to the nucleolus to the relatively 'free'
chromosome end attached to the nuclear membrane.
|
|
| A wave seems to travel along the arm from the nucleolus (top) to the nuclear membrane (bottom)? |
The B DNA form is also the configuration
that acts to minimise the amplitude of the helical proton stack in core of the DNA formed by the 2 or 3 hydrogen base-pair
bonds. These can effectively be modelled as a hydrogen crystal ribbon threading through the core of the DNA
(see illustration below).
There are naturally going to be perversities
in the operation of biological systems against the systems built by the physicists, as here in the operation of what would
seem to be a photonic crystal array. The physicists would seek to perfect the crystal structure and, therefore, the
photon/quantum/information transmission, minimising incidental losses as 'heat' to the embedding system and effectively
achieving the construction of a superconducting system. Living systems in fact depend on that precisely not being
the case. Photons as quanta or information might be something with which science can play, but living systems have
more absolute need of the photons they capture and use them to their own ends against the drive to equilibrium guaranteed
by the second law.
Metaphase
state
Moving to the other of the significant cellular states for the model, illustrated below
is the metaphase state, a minimum state for biological working, but suspected to be a maximum state for the physics
that is driving the system. That is to say, the biological working is just basal metabolism and the tail-end working
out in the cell cytoplasm of the last generation of mRNA transcripts, edits and translates. The chromosomes, at
this stage, are maximally wound self-enclosed systems and dance or 'jitter' on the metaphase plate which structure
gives a common base for the chromosomes to self-assort.
|
|
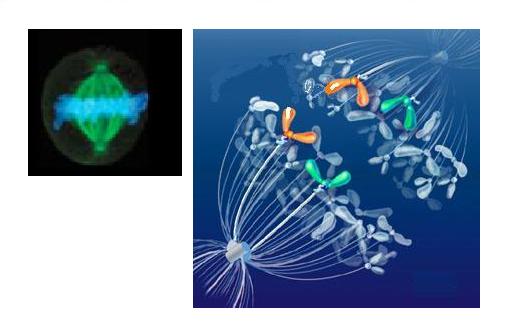
| Stained spindle (green), metaphase plate chromosomes (blue) and drawing of early separation |
At this point the DNA is maximally wound
and confined to the metaphase plate, which is a merciful reduction of the dimensional complexity for modelling purposes.
It is also then disposed, wherein the duplicated chromosome sets form the metaphase plate, as the perpendicular target
of the microtubule bundles centred at the old site of the nucleus and now occupied by the centrosome.
Whilst
for the classical biologists the working state microtubule function and description generally have to do with transport
of various molecules, for theorists such as Hameroff and Penrose, who have raised the possibility of superconduction
within these tubules, the spindle or metaphase state could offer something far more significant. If we are using
nanotubes as analogue for the possible functionalities of microtubules - and, indeed, it has been shown that stacked
microtubules, in the form of buckeytubes, allow the transport of water molecules whilst preventing the throughflow of ionic,
water-bound, atoms. In this much is raised the idea that molecular organisation can underpin an atomic
scale disequilibrium and here the spindle microtubules might be seen as feeding pure water to the chromosome or extending
the tubule entrained ion cloud to the exterior of the metaphase chromosomes.
One last consideration that perhaps needs
be mentioned here is that in the metaphase arrangements, where the chromosome set comes to form the metaphase plate, there
is maybe indication of some sort of gyroscopic functionality being realised in this formation, the 90 degrees between the
centrosomal poles and the relatively huge and now spread physical mass of the genome then acting as the gyroscopic plane.
This might make more sense later when we can pull together the more physical picture of ceullar dynamics in this respect.
Yet another interpretation of the nuclear dynamics between the working and mataphase states, in terms of the genome
mass dynamics, would be that of a bouncing ball, moving between the states of spheroidal bundle to flattened disc
...? As we will see, this at first seemingly fanciful model, in fact finds some resonance with the atomic dynamics of the
working state and we need now to move to address nuclear and genomic dynamics and reinterpret the data of biology in the light
of our needed physical understanding.
> QUANTUM LIFE
|